Chelsea M. Berns, Ph.D.
Ph.D. in Ecology and Evolutionary Biology
Worthington, A.M., C.M. Berns, and J.G. Swallow. 2012. Size matters, but so does shape: Quantifying complex shape changes in a sexually selected trait in stalk-eyed flies (Diptera:Diosidae). Biological Journal of the Linnean Society. 106: 104-113.2.
The elaborate morphologies of sexually selected ornaments are ubiquitous across the animal kingdom. In studies investigating these traits, ornament size is frequently the focus and empirical evidence supports its positive correlation with fitness. Yet, shape likely plays an important role and surprisingly, is often overlooked. Shape often changes with size, which can occur due to biomechanical, developmental, or performance constraints. Therefore, shape can provide additional insights into the morphological differences between individuals and the potential limits on sexual trait exaggeration.
In this study, we used landmark-based geometric morphometric methods to quantify head shape in a sexually dimorphic species of stalk-eyed fly (Teleopsis dalmanni) to examine patterns of sexual shape dimorphism. We also calculated univariate measurements of size (such as measurements using rulers, see Fig. 1a) and quantified overall head shape in a sexually dimorphic species of stalk-eyed fly to compare patterns of sexual shape and size dimorphism. Additionally, we investigated patterns of shape change within each sex to determine how components of head shape vary with increasing size of the eyestalks.
High resolution images of frontal views of the heads were taken in order to quantify shape and size of the eyestalks from the perspective that the flies use when competing with conspecifics and choosing mates. From these images, the locations of 40 landmarks were digitized from the images of each specimen (Fig. 1b) to represent the complex shape and curvature of the head and eyestalks.

Fig. 1a) Linear measurements of overall head morphology, and 1b) Landmarks for geometric morphometrics representing the shape of the head and eyestalks.
Our analyses revealed that stalk-eyed flies of the species Teleopsis dalmanni exhibit significant sexual dimorphism in overall head size and shape. Females have large, round eye bulbs and short, thick stalks attached to a large head, whereas males have small, elliptical eye bulbs and long, thin stalks attached to a small head (Fig. 2). These data show that the sexes differ not only in length of their eyestalks, but also in the size and shape of the individual components (i.e. eyes, stalks, and head) that compose overall head morphology. Additionally, within each sex we found a significant trend in shape change as residual eye span increased, with larger-stalked flies exhibiting a lesser angle of eyestalk attachment to the head and straighter stalks compared to those of smaller flies. Moreover, eye bulb size decreased with increasing eye span and eyestalks became thinner as they became more elongated.
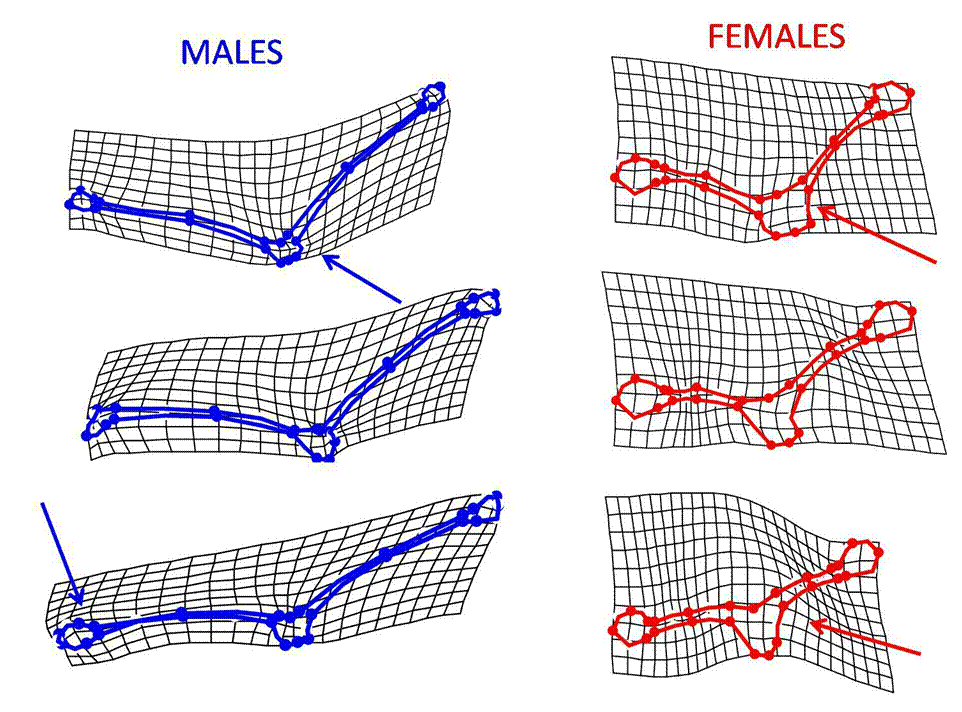
Figure 2. Thin-plate spline deformation grids visualizing change in shape with change in size for females and males with the (top) largest eye span, (middle) median eye span, and (bottom) smallest eye span. Arrows accentuate areas of the largest differences: Largest males have thinnest stalks, smallest eye bulbs and curved eyestalks, whereas the largest females have thinnest and straightest stalks with a square head. These have been accentuated by a factor of 2 to enhance interpretation.
These morphological differences within and between the sexes could be indicative of developmental constraints on eyestalk elongation. Stalk-eyed flies acquire resources during the larval phase, but the growth and maturation of adult structures occurs after pupation when feeding has ceased, thus when the exoskeleton hardens quickly, it results in structures that are fixed in size.
An alternate explanation is that of biomechanical constraints on eyestalk shape. It is reasonable to presume that large ornaments impose some costs in terms of overcoming limitations caused by gravity. The downward angle of the eyestalks attached at larger angles to the head (pointing more directly outward rather than slightly upward) may be caused by gravity acting on the eye bulbs, pulling them down prior to the exoskeleton hardening.
Patterns in the shape differences we have observed are concordant with the concept that entire suites of characters evolve to maximize whole-organism performance (Lande & Arnold, 1983; Arnold, 1983; Irschick et al., 2008). The changes in shape with increasing eye span that we observed may be correlated with flight performance, with the prediction that males will suffer from decreased aerial maneuverability without these shape changes as compared to females. Substantiating these results on performance, staged laboratory interactions with a predator indicate that males exhibit higher survival when facing a predator than do females.
Changes in ornament and head shape likely play a large role in determining moment of inertia (MOI) as well and therefore may greatly affect flight performance (Ribak et al, 2009). This may suggest that even small changes in the mass of the eye bulbs or the lateral placement of the stalks would have a significant effect on MOI, and therefore could greatly affect flight agility and maneuverability.
Our significant results demonstrate the importance of integrating geometric morphometric methods into future studies examining stalk-eyed flies. An important next step will be to expand on this analysis to include many species of stalk-eyed flies from diverse lineages to determine if these patterns of shape change are consistent across taxa.
References:
Arnold SJ. 1983. Morphology, performance, and fitness. American Zoologist 23: 347-361.
Lande R, Arnold SJ. 1983. The measurement of selection on correlated characters. Evolution 37: 1210-1226.
Ribak G, Pitts ML, Wilkinson GS, Swallow JG. 2009. Wing shape, wing size and sexual-dimorphism in eye-span in stalk-eyed flies. (Diopsidae). Biological Journal of the Linnean Society 98: 860-871.
Further Reading:Worthington, A.M., C.M. Berns, and J.G. Swallow. 2012. Size matters, but so does shape: Quantifying complex shape changes in a sexually selected trait in stalk-eyed flies (Diptera:Diosidae). Biological Journal of the Linnean Society. 106: 104-113.2.
Allen BJ, Levinton JS. 2007. Costs of bearing a sexually selected ornamental weapon in a fiddle crab. Functional Ecology 21: 154-161.
Cotton S, Rogers DW, Small J, Pomiankowski A, Fowler K. 2006. Variation in preference for a male ornament is positively associated with female eyespan in the stalk-eyed fly Diasemopsis meigenii. Proceedings of the Royal Society B 273: 1287-1292.
Evans MR, Thomas ALR. 1992. The aerodynamical and mechanical effects of elongated tails in the scarlet-tufted malachite sunbird: measuring the cost of a handicap. Animal Behavior 43:337-347.
Hingle A, Fowler K, Pomiankowski A. 2001. Size-dependent mate preference in the stalk-eyed fly Cyrtodiopsis dalmanni. Animal Behaviour 61: 589-595.
Husak JF, Ribak G, Wilkinson GS, Swallow JG. 2011. Compensation for exaggerated eyestalks in stalk-eyed flies (Diopsidae). Functional Ecology 25: 608-616.
Schillito FJ. 1971. Dimorphism in flies with stalked eyes. Zoological Journal of the Linnean Society 50:297-305.
Zahavi A. 1975. Mate selection - a selection for a handicap. Journal of Theoretical Biology 53: 205-214.